
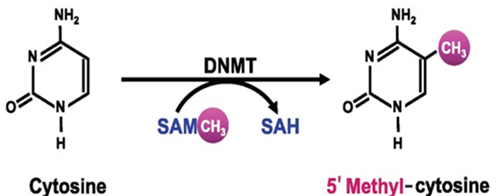
DNA由4种碱基构成,分别为胞嘧啶(C)、胸腺嘧啶(T)、腺嘌呤(A)、鸟嘌呤(G)。DNA甲基化是最早发现的基因表观修饰方式之一,真核生物中的甲基化仅发生于胞嘧啶。DNA甲基化是指由DNA甲基转移酶(DNMT)介导,在胞嘧啶的第5位碳原子上加上一个甲基基团(-CH3),使之变成5-甲基胞嘧啶(5-mC)的化学修饰过程。DNA甲基化通常抑制基因表达,去甲基化则诱导了基因的重新活化和表达。这种DNA修饰方式在不改变基因序列前提下实现对基因表达的调控。
DNA甲基化与遗传物质的稳定性
在生物个体的生长发育与繁殖过程中,维持遗传物质的稳定性是至关重要的。细胞采用多种机制来保证DNA复制的忠实性,如DNA的双螺旋结构与半保留复制模式为遗传物质的稳定提供了物质基础;DNA聚合酶Ⅲ除了具有DNA聚合酶活性外还具有5’到3’的核酸外切酶活性,可及时去除错配掺人的碱基;DNA复制后存在多种修复机制进一步保证了遗传物质的稳定性。DNA甲基化在DNA复制起始、错配修复、细菌中寄主控制的修饰与限制以及转座子的失活等过程中对维持遗传信息的稳定性发挥着重要的作用。

DNA甲基化与基因表达调控
在真核生物基因组中,基因仅仅占一小部分,例如在人类基因组中基因的编码序列还不到2%,那么在大量非编码DNA存在的情况下,实现精确控制基因的表达,降低周围的转录噪音对生物体至关重要。DNA甲基化作为一种可遗传的修饰方式为非编码DNA(内含子、重复元件以及潜在的具有活性的转座子)的长期沉默提供了一种有效的抑制机制。DNA复制后胞嘧啶的甲基化会改变DNA的构象,使DNA的大沟无法与DNA结合蛋白正常结合,从而使这些非编码区长期保持无表达活性的状态。而有转录活性的基因可利用非甲基化的启动子来进行转录表达,即使在相邻的非转录区是高度甲基化的,其启动子仍然可以起始转录并被调控。
DNA甲基化与表观遗传学
过去人们一直以为遗传和环境两大因素共同决定生物体的性状,然而人们无法合理解释马和驴的正反交后代、同卵双胞胎差别以及x染色体失活等现象。1942年Waddington首次提出了表观遗传学(epigenetics)的概念,它针对研究基因型与表型的关系,使经典的孟德尔核内遗传规律无法解释的现象得到了合理而完美的解释。基因组表观遗传修饰主要包括DNA甲基化修饰与核小体中组蛋白的修饰等,使得被修饰DNA的空间结构发生改变或使染色体结构发生改变,导致基因的沉默或过度表达。这两种修饰都是在不改变DNA碱基种类与数量的前提下使生物体表型呈现出多样化。DNA甲基化对基因表达模式以及基因组稳定性均起着至关重要的作用,并在印迹基因与x染色体失活等典型的表观遗传现象中起重要作用。

DNA甲基化与胚胎发育
DNA甲基化作为一种可遗传的表观遗传修饰,在体细胞增殖过程中通过依赖于DNA复制的DNA甲基转移酶Dnmtl稳定地传递给子细胞。但在胚胎发育的不同时期,基因组范围内的DNA甲基化水平会发生剧烈的改变,改变最剧烈的阶段为配子形成期与早期胚胎发育阶段,甲基化模式在配子形成时已经建立旧1。DNA甲基化对胚胎正常发育和等位基因的选择表达至关重要。错误甲基化模式的建立将引起人类的疾病,如Prader-Willi综合征、Angelman综合征和脆性x染色体综合征等。
